| نقشه برداری ژنتیکی ژنهای انسان بر روی کروموزومها از شیوه تجزیه و تحلیل پیوستگی ، برای تعیین فواصل بین ژنها استفاده میکند. تجزیه و تحلیل پیوستگی بر پایه اندازهگیری احتمال باقی ماندن دو ژن در کنار یکدیگر (پیوسته ماندن) در طی تقسیم میوز در گذر از یک نسل به نسل بعدی استوار است. |
دید کلی
پیوستگی را میتوان به صورت تمایل اللهای نزدیک هم ، روی یک
کروموزوم در انتقال با یکدیگر به صورت یک واحد دست نخورده در طی
میوز تعریف کرد. تجزیه و تحلیل پیوستگی ژنتیکی ، شیوهای از نقشه برداری ژنهاست که از مطالعات روی خانوادهها برای تعیین اینکه آیا دو ژن هنگام انتقال از یک نسل به نسل بعدی پیوستگی نشان میدهند (یعنی پیوستهاند) یا خیر ، استفاده میکند. نقشه برداری با تجزیه و تحلیل پیوستگی ژنتیکی با نقشه برداری به کمک شیوههای فیزیکی تفاوت دارد.
از این جهت که
نقشهبرداری فیزیکی متکی بر داشتن یک روش آزمایشگاهی جهت تعیین محل یک ژن توسط FISH یا به کمک هیبریدسازی سلولهای پیکری است. در مقابل ، تجزیه و تحلیل پیوستگی ، روش بسیار مهم و پر قدرتی در
ژنتیک پزشکی میباشد. زیرا تنها شیوهای است که نقشهبردای از ژنهای صرفا قابل شناسایی به صورت صفات فنوتیپی (شامل ژنهای بیماری) را مقدور میسازد. اکثریت ژنهای زمینهای بیماریهای ژنتیکی در این گروه قرار میگیرند، زیرا نه اساس بیوشیمیایی و نه پایه مولکولی آنها هیچ یک هنوز روشن نشده است.
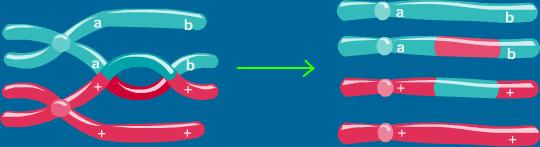
نوترکیبی هومولوگ در میوز
برای درک کامل مفاهیم تجزیه و تحلیل پیوستگی ژنتیکی ، مرور مختصر رفتار کروموزومها و ژنها در طی میوز لازم است. کروموزومهای همولوگ در طی میوز I جفت میشوند و این جفتها در طول دوک میوزی قرار میگیرند. همولوگهای پدری و مادری از طریق
تبادل متقاطع ، قطعات کروموزومی را معاوضه میکنند و کروموزومهای جدیدی را که وصله وصله (Patch work) میباشند، یکی در میان شامل بخشهایی از کروموزومهای مادر بزرگ و کروموزومهای پدربزرگ هستند، ایجاد میکنند. ایجاد این کروموزومها به مفهوم منحصر به فرد بودن ژنتیکی انسان تاکید دارد، هر
کروموزوم به ارث رسیده در فرزند یک والد ، اساسا هرگز با هیچ یک از دو نسخه کروموزوم در والد یکسان نیست.
اگر کروموزومهای همولوگ در زیر
میکروسکوپ یکسان به نظر برسند باید راهی برای افتراق همولوگ از هم داشته باشیم تا معین کنیم آیا وقایع نوترکیبی در طول کروموزومهای همولوگ رخ دادهاند و در کجا؟ برای این منظور ازنشانگرهای ژنتیکی استفاده میشود. در گذشته، ژنهایی که انواع آنزیمهای الکتروفورزی یا
آنتی ژنهای سطح سلولی را رمز گردانی میکردند، به عنوان نشانگر محسوب میشدند. به هر حال در مورد پروژه ژنوم انسانی ، شاخصهای ژنتیکی اکثرا چند شکلیهای توالی
DNA هستند که با تکثیر قطعه DNA حاوی نشانگر به
روش PCR میتوان آنها را شناسایی کرد.
نشانگرهای ژنتیکی چگونه برای نقشهبرداری پیوستگی ژنتیکی استفاده میشوند؟
فرض کنید دو جایگاه نشانگر ژنتیکی 1 و 2 با اللهای D و M روی کروموزوم پدری و اللهای m و d روی کروموزوم مادری در یک فرد وجود دارند. اگر دو جایگاه روی کروموزومهای متفاوتی واقع باشند، درمیوز مستقلا مرتب میشوند. بنابراین 4 نوع فرزند با نسبتهای مساوی وجود خواهند داشت. DM و dm که غیر نوترکیب نامیده میشوند با کروموزوم والدی اولیه تطبیق میکنند و ژنوتیپهای Dm و dM که نوترکیب نام دارند نمایانگر ترکیب جدیدی از اللها هستند که با کروموزومهای والدی تفاوت دارند. وقتی تعداد ژنوتیپهای نوترکیب و غیر نوترکیب مساوی باشد، گفته میشود که این جایگاهها غیر پیوسته هستند.
جایگاههای روی یک کروموزوم، پیوسته نیستند
فرض کنید جایگاههای 2 و 1 روی یک کروموزوم قرار دارند. ژنهای واقع روی یک کروموزوم را (
Syntenic) مینامند بدون توجه به این که با چه فاصله از هم روی کروموزوم قرار گرفته باشند. میدانیم تبادل متقاطع در مرحله چهار رشتهای میوز صورت میگیرد. اگر هیچ تبادل متقاطعی بین جایگاهها روی ندهد، صرفا ژنوتیپهای غیر نوترکیب dm و DM در فرزند دیده میشوند. یک ، دو یا بیش از دو نوترکیبی روی دهنده بین دو جایگاه ژن ، فرزندانی ایجاد میکند که 50 درصد نوترکیب و 50 درصد غیر نوترکیب هستند. اگر دو جایگاه syntenic روی یک کروموزوم آن قدر از هم دور باشند که در هر میوز حداقل یک تبادل متقاطع بین آنها صورت گیرد، ژنوتیپهای نوترکیب و غیر نوترکیب به نسبتهای مساوی در فرزندان روی خواهد داد و دو جایگاه ظاهرا غیرپیوسته به نظر میرسند، انگار که این جایگاهها روی کروموزومهای مجزا قرار دارند.
اندازه گیری فاصله ژنتیکی
فاصله ژنتیکی را بر حسب واحدهایی به نام
سانتی مورگان (CM) اندازه میگیرند و به صورت فاصله ژنتیکی که بطور متوسط در یک درصد موارد نوترکیبی روی آن رخ میدهد، تعریف میشود (سانتی مورگان 100/1 مورگان است وجه تسمیه آن نام توماس هانت مورگان است که برای اولین بار تبادل متقاطع ژنتیکی را در
مگس سرکه به نام دروزوفیلا مشاهده کرد.). برای مثال اگر در یک خانوادهای در بین فرزندان در دو جایگاه ژنی 80 درصد غیر نوترکیبی و 20 درصد نوترکیبی وجود داشته باشد، میتوان تخمین زد که فاصله بین این دو جایگاه از نظر ژنتیکی ، تقریبا 20 سانتی مورگان میباشد.
نقشههای پیوستگی ژنتیکی
نقشههای پیوستگی تعداد زیادی از جایگاههای ژنی ، حتی نقشههای پیوستگی کروموزومهای کامل ، از طریق ترکیب کردن اندازهگیریهای فاصله ژنتیکی دو جایگاه ژنی دارای پیوستگی نزدیک ایجاد شدهاند. فرض کنید دو جایگاه ژنی
B و
A با فاصلهای حدود 10 سانتی مورگان پیوسته هستند. با این اطلاعات اینک میتوانیم شروع به ساختن یک نقشه ژنتیکی برای کروموزومی که جایگاههای
B و
A روی آن هستند کنیم. میتوان جایگاههای دیگری را به نقشه پیوستگی اضافه کرد، به شرطی که فواصل آنها از جایگاههای
B و
A قابل اندازه گیری باشد.
به عنوان مثال ، جایگاه سوم
C را در نظر بگیرید که از جایگاه
A به اندازه 12 سانتی مورگان و از جایگاه
B به اندازه 5 سانتی مورگان فاصله دارد. فقط با همین اطلاعات دو نقطهای ، میتوان از طریق مشاهده ترتیب سه جایگاه ژنی را نسبت به یکدیگر تعیین کرد. توجه داشته باشید فاصله
A-C که از روی فراوانی نوترکیبی اندازهگیری میشود، کمتر از مجموع فواصل
A-B و
B-C است. این عدم همخوانی به علت این واقعیت است که تبادلات متقاطع مضاعف (یکی در فاصله
A-B و دیگری در فاصله
B-C) موجب نوترکیبی بین
C و
A نمیشوند و لذا باعث برآورد کمتر از حد واقعی فاصله بین آنها میگردند.
تجزیه و تحلیل پیوستگی چند نقطهای
روش دیگر برای تعیین ترتیب سه جایگاه ، در نظر گرفتن تمام دادهها با هم است نه هر یک از تبادلات دو نقطهای بطور جداگانه که به این روند تجزیه و تحلیل چند نقطهای میگویند. اصل تجزیه و تحلیل چند نقطهای ، تعیین ترتیب نشانگرها با به حداقل رساندن تعداد تبادلات متعدد آشکار است. به ویژه در مطالعات نقشهبرداری بسیار پیچیده ، شامل دهها جایگاه نشانگر ، تجزیه و تحلیل چند نقطهای میتواند حمایت آماری قوی در مورد صحیح بودن ترتیب خاصی از نشانگرها فراهم کند. آنگاه میتوان نقشههای ایجاد شده از طریق تجزیه و تحلیل چند نقطهای را به عنوان چارچوبهایی برای ایجاد اطلاعات تشخیصی جهت استفاده در
مشاوره ژنتیکی استفاده کرد.
با افزایش توجه متمرکز روی نقشه برداری ژنی به عنوان بخشی از پروژه ژنوم انسانی ، نقشههای پیوستگی ژنتیکی جزئی از کل ژنوم انسان با استفاده از تعدادی خانواده بزرگ سه نسلی ایجاد شده است تا فراوانی نوترکیبی در بین هزاران نشانگر جامع اقماری بسیار ریز DNA با حداکثر دقت و ظرافت ممکن اندازه گیری شود. دیگر هیچ رمز ناشناخته ژنتیکی وجود ندارد و هر هفته ژنهای جدیدی از جمله بسیاری از ژنهای مهم در
پزشکی روی
نقشه ژنتیکی انسان جای داده میشوند.
رابطه بین فواصل ژنتیکی و فیزیکی
در ابتدا براساس تعداد کیاسماهای مشاهده شده در میوز I اسپرم سازی ، کل طول ژنتیکی 23 کروموزوم در ژنوم هاپلوئید انسان حدود 3000 سانتیمورگان برآورد شد. اندازهگیری اخیر و دقیقتر حدود 4300 سانتی مورگانی ، از طریق جمع زدن تمام فواصل بین هزاران نشانگر ژنتیکی که توسط مرکز همکاری پیوستگی انسانی روی نقشه ژنتیکی انسان قرار داده شدهاند، صورت گرفته است. اگر ژنوم با طول فیزیکی هاپلوئید حدود 3x10
9 جفت باز فاصله ژنتیکی تقریبا 4300 سانتی مورگان داشته باشد، یک سانتی مورگان حدودا معادل 700 هزار جفت باز خواهد بود. جدول زیر روابط بین علامتهای
ژنتیکی سلولی ، فواصل فیزیکی و فواصل ژنتیکی را خلاصه میکند و آنها را با محتوای تقریبی ژنی ارتباط میدهد.
|
| | ژنتیک سلولی | اندازه فیزیکی | فاصله ژنتیکی | محتوای ژن
| | ژنوم هاپلوئید 23 کروموزومی | 3x109 جفت باز | 4300 سانتی مورگان | 50 هزار ژن
| | یک کروموزوم متوسط | 3x108 جفت باز | 200 سانتی مورگان | 2200 ژن
| | یک نوار کروموزومی | 3x106 جفت باز | 5 سانتی مورگان | 50 ژن
|
|
به هر حال فراوانی نوترکیبی در طول یک
کروموزوم یا سرتاسر ژنوم ثابت نیست. ضمنا نوترکیبی بین دو جایگاه نیز همیشه در
میوز افراد مونث و مذکر یکسان نیست. در نتیجه فاصله ژنتیکی (که به صورت درصد نوترکیبی در میوز اندازهگیری میشود) و فاصله فیزیکی (که به صورت جفت بازها یا نوارهای کروموزومی اندازهگیری میشود) صرفا قابل مقایسه به عنوان برآورد تقریبی ابتدایی میباشند و میتوانند سنجشهای بسیار متفاوت از فاصله بین ژنها فراهم کنند.
چشم انداز بحث
نقشه برداری از ژنهای انسان ، یکی از سریع الرشدترین حوزههای مطالعه در
ژنتیک پزشکی امروزی است. داشتن نقشه ژنی کامل انسان ، به معنای دانستن جایگاه تقریبا 50000
ژن روی
24 کروموزوم ، محلهای آنها در ارتباط با یکدیگر و فواصل بین آنهاست. این اطلاعات نقشهای بسیار باارزش هستند. نه تنها از این جهت که میتوان از آن برای تشخیص بیماریها و
مشاوره ژنتیکی استفاده کرد. بلکه به این علت که روش مستقیمی برای شناسایی ژنهای مسئول بیماریهای ژنتیکی نیز فراهم میکند.
مباحث مرتبط با عنوان