مقدمه
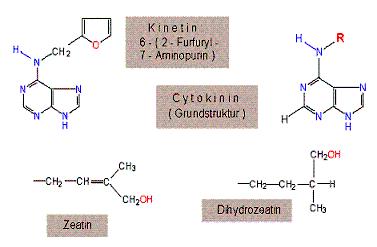
کشف سیتوکینین در سال 1965 موجب شد که گروه بسیار مهمی از تنظیم کنندههای رشد به نام سیتوکینینها مورد توجه قرار گیرد کشف قطعی سیتوکینینها در 1955 وقتی صورت گرفت که میلر و اکوگ در دانشگاه و سیکونزین مادهای به نام
کینیتین را از یک نمونه اوتوکلاو شده DNA اسپرم شاه ماهی جدا نمودند و نشان دادند که این ماده در افزایش
میتوز بافت کال توتون در شرایط آزمایشگاهی خیلی موثر است.
هابرلند در 1913 گزارش داده بوده که مواد انتشار یافته از بافت آبکشی میتواند تقسیم یاختهای را در بافت پارانشیم غدههای سیب زمینی القا کند با تکرار آزمایشهای هابرلند و ژابلوسنکی و اسکوگ در سال 1954 نشان دادند که یک قطعه از بافت آوندی کشف داده شده در روی بافت مغز توتون میتواند موجب تقسیم یاختهها مغزی که قبلا به هیچ وجه تقسیم نمیشدند گردد. این مشاهدات به یکسری تحقیقات در جهت یافتن مواد خالصی که بتوانند تقسیم یاختهای را بطور مشابه با ماده یا مواد ناشناخته موجود در بافت آوندی القا نمایند انجامید.
جداسازی کینیتین و جستجو برای یافتن سایر سیتوکینینهای طبیعی
اسکوگ و همکارانش قبلا پی برده بودند که آدنین دارای فعالیت کمی در ایجاد تقسیم یاختهای در بافت توتون است و به این موضوع اندیشیده بود که احتمالا اسیدهای نوکلئیک در فعالیت زیستی ، تحریک ناشناختهای دارند. امروزه معلوم شده است کینتین یک ترکیب مصنوعی است در گیاه وجود ندارد ولی به هر حال این ترکیب خاصیت تحریک کنندگی تقسیم یاخته را داراست و بدین ترتیب پژوهش برای ترکیبات کینتین مانند طبیعی موجود در گیاهان یعنی سیتوکینینها شدیدتر شد.
کشف سیتوکینینهای طبیعی
اولین جداسازی یک سیتوکینین طبیعی توسط لقاح از بخش میوه اوکلنونیوزلند و میلر از داشنگاه ایندیانا در حدود سال 1963 بطور همزمان انجام شد لتام در یک سمپوزیوم گزارشی ارائه کرد که در آن اعلام کرد سیتوکینین طبیعی در شکل متبلور از دانههای نارس ذرت جدا کرده است و آنرا
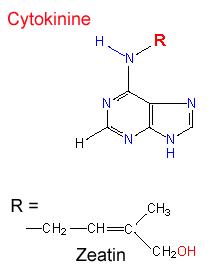
زآتین نامید. زآتین مانند کینتین ،
آدنین جانشین شده است که ساختمان آن 6- (ترانس- گاما- هیدروکسی متیل-گاما- متیل آلیل) آمینوپورین است. زنجیره جانبی آن ساختاری ترپنی دارد و در تعداد زیادی از دانهها یا میوهها مانند
آلو و
نخود و عصاره ریشهای آفتابگردان وجود دارد.
زآتین فعالترین و فروانترین سیتوکینینها باشد. بعد از کشف زآتین ، سیتوکینینهای متعدد دیگری از منابع مختلف جدا و ساختمان شیمیایی آنها مشخص گردید. تمام سیتوکینینهای طبیعی مشتقات ایزوپنتنیل آدنین (دی متیل آلیل آدنین) میباشند. سیتوکینینها در زنجیرههای بعضی از
tRNA نیز یافت میشوند که با هیدرولیز جدا میشوند. سیتوکینینها در همه گیاهان دانهدار و احتمالا در تمام قلمرو گیاهان وجود دارد.
بیوسنتز سیتوکینینها
بیوسنتز سیتوکینینها در گیاهان دانهدار عموما در بافتها و مکانهایی که مریستمی یا هنوز دارای پتانسیل رشد هستند صورت میگیرد. از ویژگیهای جالب سیتوکینینها این است که ظاهرا در ریشهها سنتز میشوند و بطور راسگرا در شاخهها انتقال مییابند. زیرا سیتوکینینها به مقدار زیادی در
شیره خام یافت شدهاند. این انتقال به طرف راس سیتوکینینها در
بافت آوندی ، در پدیده تسلط انتهایی دخالت میکند. ریشه محل عمده ، اما نه تنها محل بیوسنتز سیتوکینین است. کامبیوم و احتمالا تمام بافتهای تقسیم شونده بطور فعال مسئول سنتز سیتوکینینها میباشند.
بعضی اثرات فیزیولوژیکی سیتوکینینها در گیاهان دانهدار
سیتوکینینها در تنظیم گام به گام مراحل اوتنوژنی گیاهان دانهدار دخالت دارند از جمله:
اثرات اکسین و سیتوکینین در شکلزایی بافت کال توتون
اسکوگ طی آزمایشی به خوبی یک اثر متقابل ظریف و کمی را بین اکسین و سیتوکینین در کنترل تشکیل جوانه و ریشه از بافت مغز ساقه توتون در محیط کشت با مدرک اثبات کردند. اینها نشان دادند که با یک ترکیب خاص از غلظتهای
اکسین و کینیتین ، بافهتای مغز به صورت کال نسبتا تمایز نیافته رشد میکنند با وجود این با نسبتهای مختلف اکسین به کینتین توانستند بطور موفقی موجب تشکیل جوانه یا ریشهها از بافتهای مغز در محیط کشت شوند. نسبتهای سیتوکینین به اکسین بالا برای تشکیل جوانه و نسبتهای سیتوکینین به اکسین کم برای تشکیل ریشه مناسب بودند.
اثرات سیتوکینین در رشد و پیری
در گیاهان یکساله که چرخه زندگی در خاتمه یک فصل رویشی پایان مییابد پیری در کل گیاه مشاهده میشود. در گونههای چند ساله در انتهای یک فصل رویشی ، بخش هوایی گیاه پیر شده، میمیرد اما سیستم زیر زمینی زنده میماند. پیری یکساله و ریزش برگها در بسیاری از گیاهان چوبی چند ساله مشاهده میگردد.
پیری و مرگ معمولا از بخش قاعدهای به راسگرایی در یک شاخه پیش میرود تغییرات بیوشیمیایی که همزمان با پیری در برگها رخ میدهد بطور جامعی مشخص شدهاند مهمترین تغییرات شیمیایی در پیری کاهش در
پروتئین و
اسیدهای نوکلئیک و زردی غیر قابل برگشت در نتیجه از بین رفتن
کلروفیل است. اگر تمام برگهای جدا شده در زمان قطع کاملا جوان باشند سیتوکینین بطور محسوس رشد را تحریک میکند.
اثرات سیتوکینینها و اکسینها در تسلط راسی
تسلط راسی به بازدارندگی از رشد جوانههای جانبی زیرین توسط رشد انتهایی شاخه اطلاق میشود. مکانیسم تسلط راسی خود شناخته نشده است. این پدیده یک آنتاگونیسم بین سیتوکینینها و اکسینها را در بردارد. اکسین تولید شده در راس شاخه و انتقال یافته به پائین از آزادی جوانهها از تسلط راسی جلوگیری به عمل میآورد و این عمل اکسینها با سیتوکینینها که بعضی در خود جوانههای جانبی مهار شده سنتز میشوند و بعضی دیگر در نزدیک جوانه در شیره خام انتقال مییابد مزیت دارد.
کاربرد سیتوکینین مستقیما در جوانههای جانبی مهار شده موجب آزادی از بازدارندگی میشود. همبستگی مستقیم اثرات سیتوکینین و اکسین در مهار جوانه جانبی و رهایی از آن به خوبی اثرات این دو هورمون را در تمایز بافتهای آوندی بین جوانههای محوری و استوانه مرکزی اولیه محور اصلی شاخه به اثبات میرساند. به کار بردن اکسین روی شاخه بدون راس از تشکیل ارتباطات آوندی جلوگیری میکند و کاربرد موضعی سیتوکینین در
جوانه جانبی تمایز طرحهای آوندی را افزایش میدهد.
فعالیت هورمونی سیتوکینینهای آزاد
در مورد این نظر که سیتوکینینهای آزاد مستقلا بدون اتصال مستقیم با tRNA دارای فعالیت بیولوژیکی هستند امروزه ثابت شده است به این ترتیب که هورمون ابتدا به بعضی گیرندهها با
پیوندهای هیدروژنی یا
یونی اتصال مییابد و به نظر میرسد که گیرنده بطور عادی یک پروتئین آلوستریک باشد که در نتیجه اتصال
هورمون میتواند در اثر تغییر گیرنده یا یک کمپلکس گیرنده هورمون عمل هورمون را منجر گردد.
انتقال سیتوکینینها
در خصوص انتقال سیتوکینینها تا حدودی تناقص وجود دارد. از یک طرف اطلاعات فراوانی وجود دارد که وقتی سیتوکینینهای خارجی در برگها ، ساقهها جوانهها بکار میروند بطور قابل ملاحظهای حرکت جزئی ، اگر باشد از محل مزبور نشان میدهند. با وجود این از طرف دیگر ثابت شده است که شیره خام و
شیره پرورده به خوبی دارای سیتوکینین هستند. سیتوکینینها بطور طبیعی درشیره پرورده ، مخصوصا به صورت گلوکوزیدها وجود دارد.
احتمالا این مشاهدات ، به ظاهر مغایر واقعا غیر قابل تطبیق نیستند. سیتوکینینها در بافتهای آوندی حرکتی غیر فعال را با سایر محلولها در مطابقت صریح وابستگیهای منبع به مصرف نشان میدهند. مصرف کننده عمده برای سیتوکینینها بخشهای مریستمی یا دارای پتانسیل رویشی مثل برگهای جوان ، جوانهها ، میانگرههای جوان ، دانههای در حال نمو و میوهها میباشند.
مباحث مرتبط با عنوان