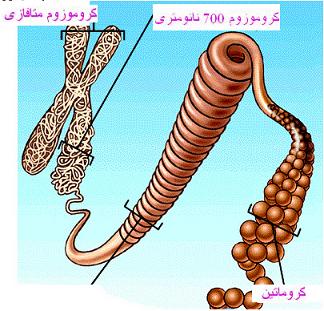
| کروموزوم در سلولهای یوکاریوتی از ترکیب کروماتین و پروتئینهای هیستونی و غیر هیستونی تشکیل شده است. و در سلولهای پروکاریوتی از ترکیب کروماتین و پروتئینهای غیر هیستونی ساخته شده است. |
نگاه کلی
واژه کروموزم به مفهوم جسم رنگی ، که در سال 1888 بوسیله
والدیر بکار گرفته شد. هم اکنون این واژه برای نامیدن رشتههای رنگپذیر و قابل مشاهده با
میکروسکوپهای نوری بکار میرود که از همانندسازی و نیز بهم پیچیدگی و تابیدگی هر رشته کروماتین اینترفازی در سلولهای یوکاریوتی تا رسیدن به ضخامت 1000 تا 1400 نانومتر ایجاد میشود. در پروکاریوتها نیز ماده ژنتیکی اغلب به حالت یک کروموزوم متراکم میشود. در برخی باکتریها علاوه بر کروموزوم اصلی که اغلب ژنها را شامل میشود کروموزوم کوچک دیگری که بطور معمول آن را پلاسمید مینامند، قابل تشخیص است گر چه تعداد کمی از ژنها بر روی پلاسمید قرار دارند.
اما از آنجا که در بیشتر موارد ژنهای مقاومت به
آنتی بیوتیکها بر روی آن جایگزین شدهاند، از نظر پایداری و بقای نسل
باکتری اهمیت زیادی دارد. کروماتین در ساختمان کروموزوم به شکل لوپ دیده میشود. لوپها توسط پروتئینهای اتصالی به
DNA که مناطق خاصی از DNA را تشخیص میدهند پابرجا میماند. سپس مراحل پیچ خوردگی نهایتا نوارهایی را که در کروموزومهای متافازی دیده میشود ایجاد میکند. هر تیپ کروموزومی یک نوع نواربندی اختصاصی را در ارتباط با نوع رنگ آمیزی نشان میدهد. این رنگ آمیزیها منجر به مشخص شدن تعداد و خصوصیات کروموزومهای هر گونه از موجودات زنده میگردد. که این خصوصیات تعدادی و مورفولوژیک کروموزومها را
کاریوتیپ مینامند.
مراحل تبدیل رشته کروماتین به کروموزوم
برای تبدیل یک رشته کروماتینی 10 تا 30 نانومتری به یک کروموزوم ، علاوه بر لزوم همانندسازی رشته کروماتین سطوح سازمان یافتگیای را در نظر میگیرند که ضمن آن با دخالت H
3 ، H
1 و پروتئینهای غیر هیستونی پیچیدگیها و تابیدگیهای رشته کروماتین افزایش مییابد، طول آن کم ، ضخامت و تراکمش زیاد میشود و به کروموزوم تبدیل میگردد. این سطوح سازمان یافتگی و اغلب به صورت رسیدن از رشته 10 تا 30 نانومتری به رشته 90 تا 100 نانومتری تشکیل رشته 30 تا 400 نانومتری و در مراحل بعد با افزایش پیچیدگیها و تابیدگیها ، ایجاد رشته 700 نانومتری و بالاخره تشکیل کروموزوم دارای دو کروماتید و با ضخامت تا 1400 نانومتر در نظر میگیرند.
اولین مرحله پیچیدگی و تراکم رشته کروماتین برای تبدیل به کروموزوم با فسفریلاسیون شدید هیستونهای H
3 ، H
1 همراه است. پس از رها شدن DNA از اکتامر هیستونی ، با دخالت آنزیمهای مسئول
همانندسازی ،
پیوندهای هیدروژنی بین دو زنجیره گسسته میشود، هر زنجیره مکممل خود را میسازد و به تدریج با ادامه همانندسازی ، دو مولکول DNA بوجود میآید که در هر مولکول یک زنجیره قدیمی و زنجیره دیگر نوساخت است. بخشهای مختلف این دو مولکول DNA که نظیر همدیگر هستند به تدریج که همانندسازیشان پایان میپذیرد، با اکتامرهای هیستونی که نیمی از آنها اکتامرهای والدی و نیمی جدید هستند ترکیب میشوند.
بعد از تشکیل ساختمان نوکلئوزومی ، دو رشته کروماتین 10 نانومتری و سپس رشتههای 30 نانومتری ایجاد میشوند. هر رشته کروماتین 30 نانومتر سطوح سازمان یافتگی را میگذارند، با مجموعهای از پروتئینهای غیر هیستونی زمینهای یا اسکلتی آمیخته میشود و به یک کروماتید تبدیل میشود. مجموعه دو کروماتید نظیر هم که از محل سانترومر بهم متصلاند
کروموزوم متافازی را ایجاد میکنند.
اجزای ساختمانی کروموزوم
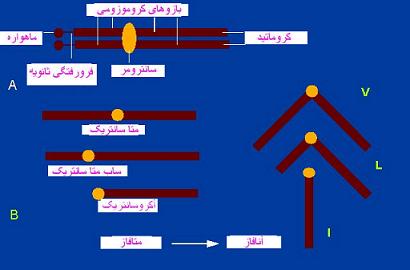
در متافاز که کروموزومها سازمان یافتگی بیشتری دارند، برای هر کروموزوم بخشهای زیر در نظر گرفته میشود.
کروماتید
کروماتید بخشی از کروموزوم متافازی است که نیمی از سراسر طول کروموزوم را میسازد. دو کروماتید هر کروموزوم از ناحیه سانترومر بهم متصلاند. هر کروماتید از ابر پیچیدگیهای رشته کروماتین و آمیختگی آن با پروتئینهای غیر هیستونی اسکلتی یا زمینهای بوجود آمده است. دو کروماتید هر کروموزوم متافازی را که در حکم تصویر آینهای یکدیگر هستند، کروماتیدهای خواهر یا کروماتیدهای نظیر مینامند.
در
پروفاز و گاهی در اینترفاز ، کروموزوم به صورت رشتههای بسیار نازکی است که آنها را کرومونما مینامند این رشتهها مراحل مقدماتی تراکم کروماتید را نشان میدهند.
کروماتید و
کرومونما ، نامی برای مشخص کردن دو ساختمان یکسان اما با دو درجه سازمان یافتگی است.
کرومومر نیز از تجمع ماده کروماتینی به صورت دانههای کروی ایجاد میشود.
سانترومر
محل اتصال دو کروماتید خواهر هر کروموزوم متافازی را سانترومر نامند. سانترومر بخش نازکی از کروموزوم که جایگاه آنرا فرورفتگی اولیه نیز مینامند. ناحیه سانترومر ناحیه بسیار هتروکروماتینی است و بویژه در بخشهای کناری خود دارای
ژنها یا ترتیبهای نوکلوتیدی تکراری است. این بخشهای هتروکروماتین با رنگهای بازی شدت رنگ میگیرند. هر کروموزوم علاوه بر سانترومر اصلی ممکن است دارای سانترومر یا سانترومرهای فرعی در محل فشردگیهای ثانویه باشد. فشردگیهای ثانویه با داشتن پیچیدگیهای کمتر از فشردگی اولیه قابل تشخیصاند.
کینه توکور
طرفین سانترمر هر کروموزوم را دو بخش پروتئینی پیاله مانند و متراکم به اسم کینه توکور میپوشاند. هر کینه توکور دارای سه بخش بیرونی و میانی و درونی است. در ساختمان هر بخش پروتئینهای رشتهای با تراکم متفاوتی قابل تشخیص هستند بخش بیرونی متراکم و بخش میانی کم تراکم است. بخش درونی بطور فشردهای با سانترومر اتصال دارد. کینه توکورها از مراکز سازماندهی
میکروتوبولها و
رشتههای دوک میتوزی هستند.
تلومر
این اصطلاح برای بخشهای انتهایی کروماتید بکار گرفته میشود. تلومرها دارای ویژگیهای سلول شناسی خاصی هستند. در
مگس سرکه ترتیبهای DNAای تلومری که در انتهای همه کروموزومها وجود دارد جدا سازی و بررسی شده است. تلومرها انتهاهای مولکولهای طویل و خطی DNAای هستند که در هر کروماتید وجود دارد. از سوی دیگر وقتی کروموزومها بوسیله عواملی مثل
پرتوهای X یا اثر آلکالوئیدها شکسته شوند، انتهاهای آزاد بدون تلومر آنها چسبنده میشود و با سایر کروموزومها ادغام میشود. علاوه بر نقشی که تلومرها در پایداری کروموزومها دارند، در برخی گونهها به حالت مهیا و بعضی بین دو کروموزوم عمل کرده و نوک به نوک اتصال موقتی پیدا میکنند.
فرورفتگی ثانویه
یکی دیگر از ویژگیهای ریخت شناسی کروموزومها هستند که از نظر موقعیت و فواصلشان بر حسب گونهها جای ثابتی دارند. وجود آنها از نظر تشخیص کروموزومها بویژه در یک مجموعه کروموزومی مفید است فرورفتگیهای ثانویه به دلیل عدم ایجاد انحرافهای زاویهدار در قطعات کروموزومی از فرورفتگیهای اولیه شناخته میشوند.
سازمان دهندگان هستکی
این نواحی فرورفتگیهای ثانویهای هستند که دارای ژنهای رمزدار کننده
RNAهای ریبوزومی جز rRNA5S میباشند و در تشکیل
هستک دخالت دارند. پدیدار شدن فرورفتگی ثانویه به دلیل رونویسی بسیار فعال ژنهای rRNAای است که آنها را از فرورفتگیهای اولیه مشخص میسازد. در انسان سازمان دهندگان هستکی در فرورفتگیهای ثانویه کروموزومهای 13 و 14 و 15 و 21 و22 قرار دارند که همه از کروموزمهای آکروسانتریک و دارای ماهواره هستند.
ماهواره
جسم کوچکی کروی است که از بقیه کروموزوم بوسیله یک فرورفتگی ثانویه جدا میشود. ماهواره و فرورفتگی ثانویه از نظر شکل و بزرگی برای هر کروموزوم ویژه ، ثابت هستند. ماهوارههای کروموزومی بخشهایی از کروموزوم از دیدگاه ریخت شناسی هستند و نبایستی آنها را با ماهوارههای DNAای که دارای ترتیبهای DNAای بسیار تکراری میباشند اشتباه کرد.
انواع کروموزمها از نظر تعداد سانترومر
کروموزومها را از نظر تعداد سانترومرهایشان به کروموزمهای یک سانترومری ، دو سانترومری و چند سانترومری تقسیم میکنند وقتی تحت تاثیر عواملی مثل
پرتوهای X کروموزمها خرد شوند و قطعاتشان ادغام شود، کروموزومهای به اصطلاح بدون سانترومر ایجاد میکنند. این کروموزومها هنگام
تقسیم سلولی رفتار عادی مثل سایر کروموزومها را ندارند.
انواع کروموزوم از نظر محل سانترومر
- کروموزمهای تلوسانتریک: سانترومر در یکی از دو انتهای کروموزومها قرار گرفته است.
- کروموزومهای آکروسانتریک: سانترومر آنها نزدیک به یکی از دو انتهای کروموزوم قرار گرفته در نتیجه یکی از بازوها نسبتا به دیگری بسیار کوچک است از قطعات کروموزومی از محل قرار گرفتن سانترومر از بازوهای کروموزومی مینامند.
- کروموزمهای متاسانتریک: سانترومر آنها در وسط کروموزوم قرار گرفته و در نتیجه بازوهای کروموزم هم اندازه هستند اکثر کروموزمها دارای یک سانترومر هستند. برخی گونهها سانترومرهای بخش شدهای دارند در رشتههای دوکی به تمامی طول کروموزوم متصلند این کروموزومها را هولوسانتریک گویند.
مباحث مرتبط با عنوان