| ژن یا ماده وراثتی (hereditary factor)، ماده پیچیدهای است که در هنگام تقسیم میتواند همانند خود را بوجود آورد. واحدهایی از این ماده وراثتی از پدر و مادر به فرزندان انتقال مییابند. این واحدها دارای ویژگیهای بسیار پایدار بوده و بطور مشخص موجودی را که صاحب آن است، تحت تاثیر قرار میدهند. ژنها بر روی کروموزومها در جایگاههای ویژه ، مرتب شدهاند. |
دید کلی
پس از آنکه
اسیدهای نوکلئیک بوجود آمدند، احتمال میرود که پیدایش جانداران جدید با سرعت بسیار زیادتری انجام گرفته باشد. این شتاب عظیم را ژنها ، که القاب کنونی اسیدهای نوکلئیک هستند امکانپذیر ساختهاند. اکنون جانداران بر طبق دستورالعملهایی که ژنهایشان فراهم میآورند، به
تولید مثل میپردازند و به سبب اینکه نسلهای متوالی جانداران ، ژنها را به ارث میبرند. پدید آمدن یک جاندار جدید به صورت فرایندی کنترل شده و غیر تصادفی درآمده است. آنچه جاندار به ارث میبرد تا حد زیادی بقای او را تعیین میکند، بنابراین وراثت از نظر سازگاری جانداران حائز اهمیت است.
اما چیزی که جانداران به ارث میبرند، ماهیچه نیرومند ، برگ سبز ، خون قرمز یا مانند آن نیست، بلکه ژنها و دیگر محتویات سلولهای زاینده است. سپس در فردی که از این سلولها ناشی میشود، صفات قابل رویت تحت نظارت ژنهایی که به ارث برده است، پدید میآید. محصول این گونه وراثت موجود زنده منحصر به فردی است که در بعضی از صفات کلی خود به والدینش شباهت دارد و در بسیاری از صفات جزئی با آنها تفاوت دارد. اگر این تفاوتها کشنده نباشند یا سبب عدم باروری نشوند، جاندار حاصل میتواند زنده بماند و ژنهای خود را به نسلهای بعدی انتقال دهد.
تاریخچه
«ویلیام هاروی» ، در سال 1651 ، این نظریه را بیان کرد که تمام موجودات زنده از جمله ، انسان ، از تخم بوجود آمدهاند و
اسپرم فقط فرایند تولید مثل نقش دارد.
هاروی همچنین تئوری اپیژنز را ارئه داد که طبق این تئوری در مرحله رشد جنینی ، ارگانها و ساختمانهای جدیدی از ماده زنده تمایز نیافته ، بوجود میآید. پژوهشهای جدید درباره وراثت بوسیله
گرگور مندل که کشیشی اتریشی بود، در نیمه دوم قرن 19 آغاز شد. وی دو قانون مهم را کشف کرد که همه پیشرفتهای بعدی علم وراثت بر پایه آنها بنا نهاده شده است.
ژن به عنوان یک واحد عملکردی
تمام نوکلئوتیدها در
DNA ، گهگاه دستخوش دگرگونیهایی میشوند که
جهش (Mutation) نام دارد. پس از هر جهش ، ژن جهش یافته (Mutant) به جای ژن اولیه به سلولهای فرزند انتقال مییابد و به ارث برده میشود. DNA جهش یافته ، آنگاه صفات تازهای بوجود میآورد که ارثی هستند. ژنهایی که جز ژنهای ساختمانی هستند، مسئول ساختن زنجیرههای پلی پپتیدی هستند.
اگر جهشی در یکی از این ژنها ، روی دهد، مجموعه صفات و ویژگیهایی که ژن جهش یافته مسئول بخش کوچکی از آن میباشد، بطور مستقیم یا غیر مستقیم ، تحت تاثیر قرار خواهند گرفت و از آنجایی که بیشتر
پروتئینها نقش آنزیمی بر عهده دارند، این جهش بر واکنشهایی که
آنزیم مربوطه در آن دخالت دارد، اثر میگذارد. ژنهای دیگر که نقش تنظیم کننده دارند، فعالیت ژنهای دیگری را کنترل میکنند و جهش در این ژنها بر کنترل ژنهای ساختمانی اثر میگذارد. DNA هر موجود از تعدادی ژنهای مختلف تشکیل شده است.
در هنگام رشد ، هر ژن دقیقا ژن همانند خود را پدید میآورد. هنگامی که یک ژن جهش مییابد، ژن جهش یافته در تقسیمات بعدی سلول ، ژنهای جهش یافته همانند خود را بوجود میآورد و اگر این ژن یک ژن ساختمانی باشد، جهش منجر به تولید پروتئین جهش یافته میگردد. ژن جهش یافته و ژن اولیه نسبت بهم آللومورف (Allelomorph) نامیده میشوند.
ژن و کروموزوم
یاختههای یک گیاه یا یک جانور دارای تعداد معینی
کروموزوم است که ویژه آن گونه گیاهی یا جانوری میباشد و تعداد این کروموزومها در همه یاختههای آن فرد پایدار و یکسان است. بنابراین همه یاختههای یک فرد دارای مجموعههای ژنی یکسانی میباشند، مثلا در
مگس سرکه در حدود 10 هزار ژن شناخته شده است. افراد مختلف یک گونه دارای
آللهای متفاوت یک ژن در سلولهای خود میباشند. در هر کروموزوم ، ژنها بطور خطی قرار گرفتهاند و نظام آنها پایدار و ثابت است. جایگاه ثابت هر ژن در کروموزوم که ویژه آن ژن است،
لوکوس (Locus) نامیده میشود.
دو ژن آلل نمیتوانند بطور همزمان در یک جایگاه وجود داشته باشند و در یک زمان هر جایگاه میتواند پذیرایی تنها یکی از ژنهای آلل باشد. برخی از ژنها به ویژه ژنهایی که در ساختن RNA دخالت دارند، چندین بار در یک مجموعه کروموزومی تکرار میشوند. در پدیده
میتوز ، پیش از تقسیم هسته ، ژنها و در نتیجه کرومزومها، دو برابر شدهاند و هر یک از دو یاخته حاصل از تقسیم ، یکی از مجموعههای کروموزومی را دریافت میکند و از اینرو مجموعههای کروموزومی دو سلول دقیقا یکسان خواهد بود.
ژن و گوناگونی افراد
در یاختههای بدنی گیاهان و جانوران کروموزومها به صورت جفت وجود دارند و از نظر ظاهری یکسان میباشند (به جز کروموزومهای جنسی). در هر لنگه از یک جفت کروموزوم ، نظام جایگاههای ژنی ، همانند نظام جایگاههای لنگه دیگر میباشد و ژنهایی که در جایگاههایی همانند قرار دارند، ممکن است یکسان بوده و یا آلل یکدیگر باشند. در حالت نخست فرد از نظر دو ژن
هموزیگوت و در حالت دوم
هتروزیگوت میباشد. شماره کروموزومها در یاختههای حاصل از
تقسیم میوز یا
گامتها ، 2/1 تعداد کروموزومها در سلولهای پیکری است و در هر یک از گامتها ، تنها یک لنگه از یک جفت کروموزوم همانند ، در برخی از جایگاهها باهم متفاوت هستند.
در نتیجه گامتها نیز با هم متفاوت خواهند بود و چون توزیع کروموزومها در هر گامت از قانون احتمالات پیروی میکند، در نتیجه احتمال تولید گامتهای مختلف در صورتی که تعداد کروموزومها را
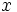
در نظر بگیریم،
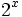
خواهد بود. این حالت ،
تفکیک مستقل نامیده میشود. تقاطع کروموزومی (Crossing-Over) نیز به ایجاد تفاوتهای بیشتر بین گامتها ، کمک میکند.
سازمان یابی و ساختمان ژن
در سادهترین حالت ، یک ژن را میتوان به صورت قطعهای از یک مولکول DNA و حاوی رمز برای توالی
اسید آمینهای یک رشته پلی پپتیدی و توالیهای تنظیم کننده لازم برای بروز آن در نظر گرفت. به هر حال این توصیف برای ژنهای موجود در
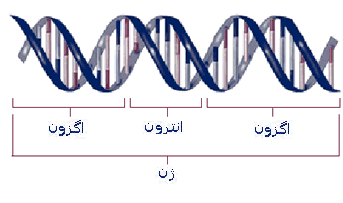
ژنوم انسان ، ناکافی است، زیرا تعداد ناچیزی ژن به صورت توالیهای رمزدار پیوسته وجود دارد. بلکه در عوض در بین اکثریت ژنها ، یک یا بیش از یک ناحیه فاقد رمز موجود است. این توالیهای حد فاصل که اینترون (intron) نامیده میشوند، ابتدا در هسته به RNA رونویسی میشوند، اما در RNA پیامبر بالغ در سیتوپلاسم وجود ندارند.
لذا اطلاعات توالیهای اینترونی ، بطور طبیعی در فرآورده پروتئینی نهائی نمایانده نمیشود. اینترونها یک در میان با توالیهای رمزدار یا
اگزون (exon) که نهایتا توالی اسید آمینهای پروتئین را رمز گردانی میکنند، قرار دارند. اگرچه تعداد کمی از ژنها در ژنوم انسان فاقد اینترون میباشند، اکثر ژنها حداقل یک و معمولا چندین اینترون دارند. ژن دیستروفین وابسته به جنس که حاوی 2 میلیون جفت باز است، کمتر از یک درصد آن حاوی اگزونهای رمزدار است. اینترونها در ساختار ژنها ، نقش حفاظت از اگزونها را در برابر جهشها بر عهده دارند.
خصوصیات ساختمانی یک ژن معمولی انسان
ژن نه تنها توالیهای رمزدار واقعی است، بلکه دارای توالیهای نوکلئوتیدی مجاور لازم برای بروز مناسب ژن ، یعنی برای تولید یک
مولکول RNA پیامبر طبیعی ، به مقدار صحیح ، در محل درست و در زمان صحیح حین تکامل و یا در طی چرخه سلولی نیز میباشد. توالیهای نوکلئوتیدی مجاور ، پیامهای مولکولی شروع و پایان را برای ساخت
RNA پیامبر رونویسی شده از ژن فراهم میکنند. ژن دارای دو انتهای
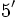
به
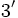
است. در انتهای
ژن ، یک ناحیه پیشبر وجود دارد که شامل توالیهای مسئول شروع مناسب رونویسی است.
پیشبرها و نیز عناصر تنظیم کننده میتوانند محلهایی برای جهش در
بیماریهای ژنتیکی که قادرند مانع بروز طبیعی ژن شوند، باشند. این عناصر تنظیم کننده شامل تقویت کنندهها ، خاموش کنندهها و نواحی کنترل کننده جایگاه ژنی هستند. در انتهای
ژن ، یک ناحیه ترجمه نشده مهم یافت میشود که حاوی پیامی برای اضافه شدن یک توالی از واحدهای آدنوزین به اصطلاح دم پلی A به انتهای RNA پیامبر بالغ است.
مبانی بروز ژن
جریان اطلاعات از ژن به
پلی پپتید ، شامل چندین مرحله است.
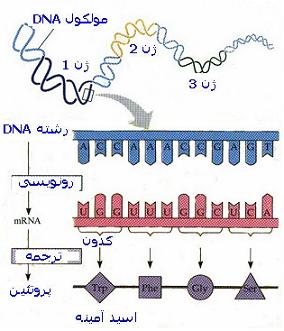
- رونویسی یک ژن در محل شروع رونویسی روی RNA کروموزومی ، بلافاصله از توالیهای رمزدار آغاز میشود و در طول کروموزوم ادامه یافته، از چند صد جفت باز تا بیش از یک میلیون جفت باز و در هر دو گروه اینترونها و اگزونها و ناحیه بعد از پایان توالیهای رمزدار را رونویسی میکند.
- پس از تغییر یافتن در هر دو انتهای و رونوشت اولیه RNA ، بخشهای مربوط به اینترونها برداشته میشوند و قطعات مربوط به اگزونها به یکدیگر چسبانده میشوند.
- پس از برش و چسباندن RNA ، RNA پیامبر حاصل که اینک فقط حاوی بخشهای رمزدار ژن است، از هسته به سیتوپلاسم سلول برده میشود و در آنجا نهایتا به توالی اسید آمینهای پلی پپتید رمزگردانی شده ، ترجمه میگردد. هر یک از این مراحل ، در معرض بروز خطا هستند و جهشهایی که در هر یک از این مراحل مداخله میکنند، در ایجاد تعدادی از اختلالات ژنتیکی دخیل دانسته شدهاند.
مباحث مرتبط با عنوان